 Reviews
Reviews
About Crinoids
Crinoids are commonly known as sea lilies, though they are animals, not plants. Crinoids are echinoderms related to starfish, sea urchins, and brittle stars. Many crinoid traits are like other members of their phylum. Such traits include tube feet, radial symmetry, a water vascular system, and appendages in multiples of five (pentameral).
Most of the Paleozoic crinoid species attached themselves to substrates on the ocean floor. Crinoids are famous for their feathery, tentacle-like appendages that opened up like a flower and captured particles of food such as plankton. Though crinoids appeared in the Ordovician (488 mya), they survived the Permian mass extinction and diversified into hundreds of species which survive, today.
Crinoid Quick Facts:
Crinoids have pentaradial, or five-fold symmetry.
Crinoidea is derived from “krinon”, which is Greek for “a lily.”
Crinoids capture food with tube feet when prey and detritus float through its feathery arms.
The largest fossil crinoid stem which has ever been found is 130 ft (40 m) long.
Crinoids once carpeted the ocean floor, swaying with the currents.
Crinoids are some of the Earth’s earliest animals. They have managed to survive and evolve through several global climate changes and mass extinctions.
Fossilized segments from the stalks have been used as beads for necklaces and rosaries.
The earliest known crinoids are from the Ordovician, which began about 488 mya. Some scientists assert that a fossil from the Cambrian Burgess Shale may record an earlier emergence. This fossil is called Echmatocrinus. The Burgess Shale is considered 500 mya. Approximately 625 species of crinoids still survive today. They are the descendants of the crinoids which survived the mass extinction at the end of the Permian. It is estimated that over 6000 species of crinoids have lived on the Earth.

A modern day stalked crinoid in the Gulf of Mexico. Photo from NOAA library.
Scientists suggest two possibilities for the origin of crinoids. The first hypothesis suggests that crinoids evolved from the blastozoan eocrinoids and cystoids. Eocrinoids were from the Early Cambrian and were the earliest echinoderms with a stalk, and arms. The group Gogiida is often referenced and is recognized by its vase-shaped body. Cystoids had stems and were similar to crinoids, but had 3 ambulacral areas on the outside of an ovoid body. A difference between the anatomy of crinoids and blastoids is that blastoids have a flower bud-shaped compartment that holds the vital organs and is called a theca, rather than a calyx. A crinoid calyx is cup-shaped. The calcareous plates that protect the outside of the theca and calyx are often referred to as thecal plates. These plates are used to identify species. The term is also used to describe dinoflagellate anatomy.
Another hypothesis is that crinoids evolved from edrioasteroids. Edrioasteroidea may have emerged as early as the Ediacaran. These organisms had a theca and 5 ambulacral arms in the body wall. It is difficult to define the exact origin for the crinoids because all possible ancestral groups share key traits such as directly attaching to substrates. Other traits include radial symmetry and calcareous plates.
The Ordovician was the first period that crinoids experienced adaptive radiation. The second period of adaptive radiation occurred in the Triassic, following the Permian mass extinction (250 mya). Flexible arms and motility became widespread in the Triassic. The traits are believed to have evolved due to pressure from predators such as benthic echinoids.
Crinoids were not the only successful stalked and filter-feeding echinoderms of the Paleozoic. Blastoids were also highly diverse and abundant. Crinoids were their most diverse in the Paleozoic, peaking in abundance and diversity in the Mississippian. Blastoids, however well suited to the Paleozoic, did not survive through the end of the Permian. Crinoids evolved new survival strategies for the environments of the Mesozoic.
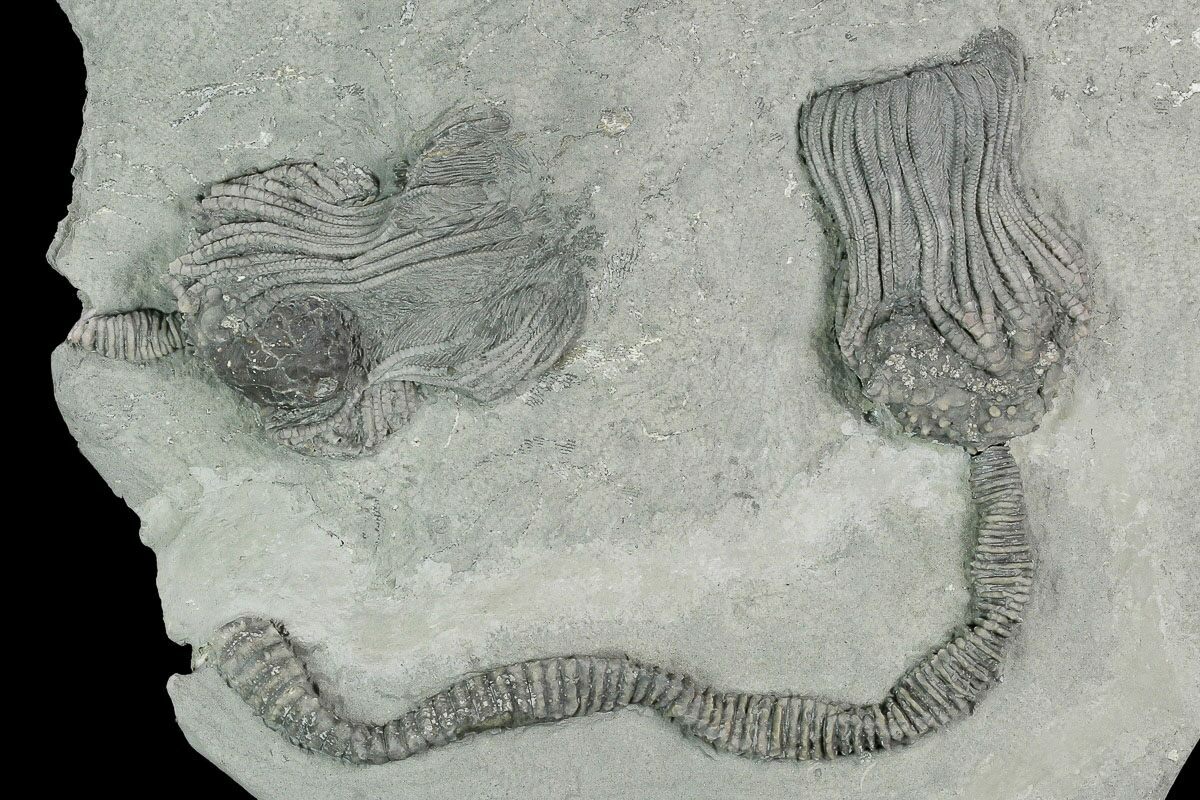
Crinoids have the pentaradial symmetry that is a characteristic of echinoderms. The three main sections of a crinoid give it the lily-like appearance. These sections are the segmented column or stem, the calyx where the body cavity and digestion occurs, and the arms which filter food from the environment. Most crinoids live attached to substrate, though there are free swimming species in the fossil record. There are several crinoid species alive today which swim freely as adults.

A fossil of a typical crinoid, showing (from bottom to top) the stem, calyx, and arms with cirri
Crinoids can very basically be described as upside-down starfish with a stems. The stem of a crinoid extends down from what would be the top of a starfish, leaving the mouth of the organism opening skyward, with the arms splayed out. However, crinoid arms look articulated and feathery. The stalk extends down from the aboral surface of the calyx. The stalk column has holdfasts which attach the animal to substrate. A holdfast does just as its name implies. Many living species have a vestigial stalk.
Stalked crinoids have been observed moving, but not on ocean currents. In 2005, a living species of stalked crinoid near Grand Bahama Island was observed moving itself across the sea floor. Before the recording, the fastest motion on record was 2 ft/hr, though some scientists think that faster speeds are possible. Paleozoic crinoids were stationary. Motility developed later in response to predation.
The crinoid column is also called a stem or stalk. The stem is made from disc-shaped pieces of endoskeleton which are stacked upon each other and are hollow in the middle. They are held together by ligaments which decomposed rapidly after death. The hollow shapes inside of the discs include: elliptical, circular, pentagonal, and star-shaped. Column discs which are square-shaped have 5 holes in them. Fossil discs are commonly found in Paleozoic limestone deposits.
The stem is constructed from a biomineral complex made from calcium carbonate crystals in a sponge-like microstructure. The three-dimensional microstructure is called stereom. In the embryo, the stereom is produced by mesenchymal cells, which makes stereom an endoskeletal feature of echinoderms. The calcium carbonate crystals which are aligned in the stereom structure, plus the stereom lattice, together create what are called ossicles. Sclerocytes excrete ossicles. Ossicles are held together by collagenous ligaments and are covered by epidermis. The ligaments provide a variety of postures for the organism without using extra muscular energy. Though covered by tissue, the ossicles of echinoderms have the function of an exoskeleton.
Some species have special appendages emerging from the bottom of the calyx or along the stem. These appendages are not used for feeding, but are types of holdfasts. They are called cirri. Cirri are used for attaching crinoids to different things, perhaps even to other crinoids. Cirri can be embedded into mud to stabilize the column. Cirri have different characteristics depending on the species. Cirri can act as a hook, or wrap around an object. Another type of holdfast is a button-like place on the bottom of the stem that can strongly adhere to substrate such as coral.

A fossilized holdfast of the crinoid Eucalyptocrinites which would have anchored it to the substrate.
The arms, or brachials, of the crinoid can splay open for filter feeding. There are typically five arms found on the fossil types, though some living species can have many more arms- always in multiples of five. Smaller ossicles form the structure of the arms. The arms of the crinoid have tube feet and cilia on them which move food along the ambulacral canal of each arm. From each canal, food is transported by cilia into the mouth. The arms and calyx are called the crown.
The calyx is a cup-shaped body part which contains the u-shaped digestive system and the organs for reproduction. Five sided, calcareous plates create the lower cup of the calyx. The plates are in rows of five and radiate around the calyx. The mouth is atop the dorsal cup, and the anus is beside it. The tegmen is the skyward surface of the calyx. The tegmen has five ambulacral areas which includes a deep groove in each area. Arms extend from the grooved areas. Tube feet are arranged along pinnules which project from the jointed arms. The visual effect makes the arms appear feathery and allows the crinoid to comb the water for suspended food. The hydraulic pressure in each tube foot is controlled with a water vascular system connected to the body cavity.
The body cavity consists of connective tissue and narrow canals which extend into the arms and column. The haemal system of crinoids is a network of fluid filled sinuses. These sinuses are placed among the connective tissue. A group of sinuses surround the oesophogus. The fluid filled spaces extend toward the bottom of the calyx where glandular tissue is located. These spaces are where nutrients are transported to cells of the body. The sinuses also act as an excretory and respiratory system. Tube feet are thinly walled and allow oxygen to be absorbed by crinoids. Coelomocytes are phagocytic cells and are responsible for capturing waste.
The tube feet which project from the pinnules are coated with sticky mucus. The mucus captures particles of food that floats by. Once a food particle is captured, the tube feet move the particle into the ambulacral groove. From inside the groove, cilia move mucus and the particle toward the mouth.
There is an observed correlation, in some living species, with the number of branches on the arms pertaining to the abundance of food in the habitat. The more rich the food supply, the fewer the number of arms needed to obtain food. Ancient crinoids typically had 5 arms, though living species predominantly have arms which divide into two branches, which results in 10 arms.
Crinoids do not have a stomach, so food particles go from mouth to short esophagus and then to intestine. The intestine makes a single curve on the inside of the calyx and ends with a rectum and an anus at the margin of the tegmen.
Crinoids consumed bits of matter and small organisms. Nutrients were obtained from diatoms, plankton, detritus, larva, and anything that was the appropriate size to be trapped by the pinnules and moved into the ambulacral canal.
The nervous system of a crinoid consists of a nerve ring around the mouth with nerves that extend to each arm. There is another ring with branched nerves to each arm that is associated with senses, and a third network with a neural mass at the base of the calyx with extensions to each arm and the stalk. The third nerve network controls motor action.
Crinoids are male or female (dioecious). Crinoids have genital canals that produce gametes, but they do not have gonads. The canals are in some of the pinnules which break open to release sperm and eggs. The fertilized eggs eventually hatch in the water. The larva is barrel-shaped with ciliated bands and sensory hairs.. These are known as vitellaria larvae. The larva settles and attaches to substrate with an adhesive gland. Metamorphosis begins and the larva becomes a stalked adult. Crinoids living today can reproduce in about 10 to 16 months, and even the free-swimming species sometimes mature on the stalk before breaking away.
Crinoids were very successful in the Paleozoic. They were most abundant and diverse in the Mississippian when several continents were covered with shallow seas. Thick layers of limestone stratigraphy throughout North America and Europe are full of crinoid stems.
In the Ordovician, which began about 488 mya, sea levels were at an all-time high. The globe north of the equator was almost completely ocean. The supercontinent Gondwana was oriented in the southern hemisphere and it contained most of the land. The Taconic Orogeny is a notable event which occurred on a subduction zone as a volcanic island arc collided with the North American continent. The remnants of this collision are seen in eastern New York and Connecticut. The orogeny waned to conclusion in the Silurian. This event is significant to crinoid preservation. The erosion of the sediments produced by the orogeny supplied the surrounding sea with mud and clay which preserved fauna during bursts of undersea mudslides and rapid storm deposition.
Between 450 mya and 440 mya, there were two periods of extinction that together form the second largest mass extinction that has occurred on the planet. Triggers included the tectonic movement of Gondwana which had moved to the south polar region. A drop in CO2 levels also occurred. The result was a cooling of the planet which allowed the formation of glaciers. The glaciations then caused periodic drops in sea levels which adversely affected life living on the continental shelves.
The Silurian began about 444 mya. It was a warmer period, with fewer glaciations and higher sea levels, though glaciers still persisted on a united Gondwana in the southern hemisphere. The supercontinent Euramerica formed at the equator. Jawed and bony fish appeared in the seas, while vascular plants emerged on land. The climate was warmer and more stable than in the Ordovician.
The Devonian began 416-419 mya. In the Devonian, Euramerica and Gondwana became closer together and the climate was warm. Sea levels were high and shallow reefs and coral communities were abundant. The Devonian is known as the Age of Fish.
Plate tectonics continued to alter the continental landscape and crinoids continued to diversify and populate ocean habitats throughout the Carboniferous. The supercontinent Pangaea was along the equator in the Permian (299-250 mya).
At the beginning of the Permian, sea levels were relatively low and shallow marine habitats would have been greatly reduced. The climate was warming, though. As a result, glaciers that were present at the beginning of the Permian had disappeared by its end. The enormous landmasses promoted extreme variations in climate. At the end of the Permian, 90 percent of life in the sea had gone extinct. Crinoids emerged from the extinction as a few surviving groups. They adapted and diversified, and their descendants survive today.
Articulated fossils are found where currents were too weak to scatter the calcareous plates and discs, and burial occurred before bacteria could break down the ligaments. The Middle Ordovician layers of the Trenton group are known for excellent specimens. This is due to a period of colonization by crinoids when the deposition of sediment was low. An event such as a storm created a massive deposit of fine-grained mud which eventually slid and rapidly buried the colonies, preserving structures in place.
Orange and yellow organic molecules were discovered by C. O’Malley at Ohio State University. The molecules were found in 350 million year old crinoids from the Mississippian period.
In 2003, Shibata and Oji reported that at least some living crinoids can intentionally drop an arm as they mature, enabling it to have the full number of arms for its adult form. The arm is then replaced by two arms. The animal can repeat the process until it is mature.
Marsupitsa, Saccocoma, and Uintachrinus are considered free-swimming. Uintacrinus was a stalk-less crinoid with long arms for capturing prey. It was free-swimming the Western Interior Seaway of Cretaceous North America, 70 mya.
Pentacrinites is a fossil species which attached to driftwood. Pentacrinites evolved the ability to grow lengthy stems, even several meters long. This was likely a response to the sinking wood.
Delocrinus missouriensis is the state fossil of Missouri. It is found in Pennsylvanian period deposits.
Ancyrocrinus is found in Devonian sediments. Its cirri had evolved into a grappling hook.
Himerocrinus has been found with more than 80 arms.
Most of the Paleozoic crinoid species attached themselves to substrates on the ocean floor. Crinoids are famous for their feathery, tentacle-like appendages that opened up like a flower and captured particles of food such as plankton. Though crinoids appeared in the Ordovician (488 mya), they survived the Permian mass extinction and diversified into hundreds of species which survive, today.
Crinoid Quick Facts:
Taxonomy and Origins
The earliest known crinoids are from the Ordovician, which began about 488 mya. Some scientists assert that a fossil from the Cambrian Burgess Shale may record an earlier emergence. This fossil is called Echmatocrinus. The Burgess Shale is considered 500 mya. Approximately 625 species of crinoids still survive today. They are the descendants of the crinoids which survived the mass extinction at the end of the Permian. It is estimated that over 6000 species of crinoids have lived on the Earth.

A modern day stalked crinoid in the Gulf of Mexico. Photo from NOAA library.
Scientists suggest two possibilities for the origin of crinoids. The first hypothesis suggests that crinoids evolved from the blastozoan eocrinoids and cystoids. Eocrinoids were from the Early Cambrian and were the earliest echinoderms with a stalk, and arms. The group Gogiida is often referenced and is recognized by its vase-shaped body. Cystoids had stems and were similar to crinoids, but had 3 ambulacral areas on the outside of an ovoid body. A difference between the anatomy of crinoids and blastoids is that blastoids have a flower bud-shaped compartment that holds the vital organs and is called a theca, rather than a calyx. A crinoid calyx is cup-shaped. The calcareous plates that protect the outside of the theca and calyx are often referred to as thecal plates. These plates are used to identify species. The term is also used to describe dinoflagellate anatomy.
Another hypothesis is that crinoids evolved from edrioasteroids. Edrioasteroidea may have emerged as early as the Ediacaran. These organisms had a theca and 5 ambulacral arms in the body wall. It is difficult to define the exact origin for the crinoids because all possible ancestral groups share key traits such as directly attaching to substrates. Other traits include radial symmetry and calcareous plates.
The Ordovician was the first period that crinoids experienced adaptive radiation. The second period of adaptive radiation occurred in the Triassic, following the Permian mass extinction (250 mya). Flexible arms and motility became widespread in the Triassic. The traits are believed to have evolved due to pressure from predators such as benthic echinoids.
Crinoids were not the only successful stalked and filter-feeding echinoderms of the Paleozoic. Blastoids were also highly diverse and abundant. Crinoids were their most diverse in the Paleozoic, peaking in abundance and diversity in the Mississippian. Blastoids, however well suited to the Paleozoic, did not survive through the end of the Permian. Crinoids evolved new survival strategies for the environments of the Mesozoic.
Anatomy
Crinoids have the pentaradial symmetry that is a characteristic of echinoderms. The three main sections of a crinoid give it the lily-like appearance. These sections are the segmented column or stem, the calyx where the body cavity and digestion occurs, and the arms which filter food from the environment. Most crinoids live attached to substrate, though there are free swimming species in the fossil record. There are several crinoid species alive today which swim freely as adults.

A fossil of a typical crinoid, showing (from bottom to top) the stem, calyx, and arms with cirri
Crinoids can very basically be described as upside-down starfish with a stems. The stem of a crinoid extends down from what would be the top of a starfish, leaving the mouth of the organism opening skyward, with the arms splayed out. However, crinoid arms look articulated and feathery. The stalk extends down from the aboral surface of the calyx. The stalk column has holdfasts which attach the animal to substrate. A holdfast does just as its name implies. Many living species have a vestigial stalk.
Stalked crinoids have been observed moving, but not on ocean currents. In 2005, a living species of stalked crinoid near Grand Bahama Island was observed moving itself across the sea floor. Before the recording, the fastest motion on record was 2 ft/hr, though some scientists think that faster speeds are possible. Paleozoic crinoids were stationary. Motility developed later in response to predation.
Stem
The crinoid column is also called a stem or stalk. The stem is made from disc-shaped pieces of endoskeleton which are stacked upon each other and are hollow in the middle. They are held together by ligaments which decomposed rapidly after death. The hollow shapes inside of the discs include: elliptical, circular, pentagonal, and star-shaped. Column discs which are square-shaped have 5 holes in them. Fossil discs are commonly found in Paleozoic limestone deposits.
The stem is constructed from a biomineral complex made from calcium carbonate crystals in a sponge-like microstructure. The three-dimensional microstructure is called stereom. In the embryo, the stereom is produced by mesenchymal cells, which makes stereom an endoskeletal feature of echinoderms. The calcium carbonate crystals which are aligned in the stereom structure, plus the stereom lattice, together create what are called ossicles. Sclerocytes excrete ossicles. Ossicles are held together by collagenous ligaments and are covered by epidermis. The ligaments provide a variety of postures for the organism without using extra muscular energy. Though covered by tissue, the ossicles of echinoderms have the function of an exoskeleton.
Appendages
Some species have special appendages emerging from the bottom of the calyx or along the stem. These appendages are not used for feeding, but are types of holdfasts. They are called cirri. Cirri are used for attaching crinoids to different things, perhaps even to other crinoids. Cirri can be embedded into mud to stabilize the column. Cirri have different characteristics depending on the species. Cirri can act as a hook, or wrap around an object. Another type of holdfast is a button-like place on the bottom of the stem that can strongly adhere to substrate such as coral.

A fossilized holdfast of the crinoid Eucalyptocrinites which would have anchored it to the substrate.
The arms, or brachials, of the crinoid can splay open for filter feeding. There are typically five arms found on the fossil types, though some living species can have many more arms- always in multiples of five. Smaller ossicles form the structure of the arms. The arms of the crinoid have tube feet and cilia on them which move food along the ambulacral canal of each arm. From each canal, food is transported by cilia into the mouth. The arms and calyx are called the crown.
Calyx
The calyx is a cup-shaped body part which contains the u-shaped digestive system and the organs for reproduction. Five sided, calcareous plates create the lower cup of the calyx. The plates are in rows of five and radiate around the calyx. The mouth is atop the dorsal cup, and the anus is beside it. The tegmen is the skyward surface of the calyx. The tegmen has five ambulacral areas which includes a deep groove in each area. Arms extend from the grooved areas. Tube feet are arranged along pinnules which project from the jointed arms. The visual effect makes the arms appear feathery and allows the crinoid to comb the water for suspended food. The hydraulic pressure in each tube foot is controlled with a water vascular system connected to the body cavity.

The body cavity consists of connective tissue and narrow canals which extend into the arms and column. The haemal system of crinoids is a network of fluid filled sinuses. These sinuses are placed among the connective tissue. A group of sinuses surround the oesophogus. The fluid filled spaces extend toward the bottom of the calyx where glandular tissue is located. These spaces are where nutrients are transported to cells of the body. The sinuses also act as an excretory and respiratory system. Tube feet are thinly walled and allow oxygen to be absorbed by crinoids. Coelomocytes are phagocytic cells and are responsible for capturing waste.
Feeding and Diet
The tube feet which project from the pinnules are coated with sticky mucus. The mucus captures particles of food that floats by. Once a food particle is captured, the tube feet move the particle into the ambulacral groove. From inside the groove, cilia move mucus and the particle toward the mouth.
There is an observed correlation, in some living species, with the number of branches on the arms pertaining to the abundance of food in the habitat. The more rich the food supply, the fewer the number of arms needed to obtain food. Ancient crinoids typically had 5 arms, though living species predominantly have arms which divide into two branches, which results in 10 arms.
Crinoids do not have a stomach, so food particles go from mouth to short esophagus and then to intestine. The intestine makes a single curve on the inside of the calyx and ends with a rectum and an anus at the margin of the tegmen.
Crinoids consumed bits of matter and small organisms. Nutrients were obtained from diatoms, plankton, detritus, larva, and anything that was the appropriate size to be trapped by the pinnules and moved into the ambulacral canal.
Nervous System
The nervous system of a crinoid consists of a nerve ring around the mouth with nerves that extend to each arm. There is another ring with branched nerves to each arm that is associated with senses, and a third network with a neural mass at the base of the calyx with extensions to each arm and the stalk. The third nerve network controls motor action.
Reproduction
Crinoids are male or female (dioecious). Crinoids have genital canals that produce gametes, but they do not have gonads. The canals are in some of the pinnules which break open to release sperm and eggs. The fertilized eggs eventually hatch in the water. The larva is barrel-shaped with ciliated bands and sensory hairs.. These are known as vitellaria larvae. The larva settles and attaches to substrate with an adhesive gland. Metamorphosis begins and the larva becomes a stalked adult. Crinoids living today can reproduce in about 10 to 16 months, and even the free-swimming species sometimes mature on the stalk before breaking away.
Environment
Crinoids were very successful in the Paleozoic. They were most abundant and diverse in the Mississippian when several continents were covered with shallow seas. Thick layers of limestone stratigraphy throughout North America and Europe are full of crinoid stems.
In the Ordovician, which began about 488 mya, sea levels were at an all-time high. The globe north of the equator was almost completely ocean. The supercontinent Gondwana was oriented in the southern hemisphere and it contained most of the land. The Taconic Orogeny is a notable event which occurred on a subduction zone as a volcanic island arc collided with the North American continent. The remnants of this collision are seen in eastern New York and Connecticut. The orogeny waned to conclusion in the Silurian. This event is significant to crinoid preservation. The erosion of the sediments produced by the orogeny supplied the surrounding sea with mud and clay which preserved fauna during bursts of undersea mudslides and rapid storm deposition.
Between 450 mya and 440 mya, there were two periods of extinction that together form the second largest mass extinction that has occurred on the planet. Triggers included the tectonic movement of Gondwana which had moved to the south polar region. A drop in CO2 levels also occurred. The result was a cooling of the planet which allowed the formation of glaciers. The glaciations then caused periodic drops in sea levels which adversely affected life living on the continental shelves.
The Silurian began about 444 mya. It was a warmer period, with fewer glaciations and higher sea levels, though glaciers still persisted on a united Gondwana in the southern hemisphere. The supercontinent Euramerica formed at the equator. Jawed and bony fish appeared in the seas, while vascular plants emerged on land. The climate was warmer and more stable than in the Ordovician.
The Devonian began 416-419 mya. In the Devonian, Euramerica and Gondwana became closer together and the climate was warm. Sea levels were high and shallow reefs and coral communities were abundant. The Devonian is known as the Age of Fish.
Plate tectonics continued to alter the continental landscape and crinoids continued to diversify and populate ocean habitats throughout the Carboniferous. The supercontinent Pangaea was along the equator in the Permian (299-250 mya).
At the beginning of the Permian, sea levels were relatively low and shallow marine habitats would have been greatly reduced. The climate was warming, though. As a result, glaciers that were present at the beginning of the Permian had disappeared by its end. The enormous landmasses promoted extreme variations in climate. At the end of the Permian, 90 percent of life in the sea had gone extinct. Crinoids emerged from the extinction as a few surviving groups. They adapted and diversified, and their descendants survive today.
Discoveries
Articulated fossils are found where currents were too weak to scatter the calcareous plates and discs, and burial occurred before bacteria could break down the ligaments. The Middle Ordovician layers of the Trenton group are known for excellent specimens. This is due to a period of colonization by crinoids when the deposition of sediment was low. An event such as a storm created a massive deposit of fine-grained mud which eventually slid and rapidly buried the colonies, preserving structures in place.
Orange and yellow organic molecules were discovered by C. O’Malley at Ohio State University. The molecules were found in 350 million year old crinoids from the Mississippian period.
In 2003, Shibata and Oji reported that at least some living crinoids can intentionally drop an arm as they mature, enabling it to have the full number of arms for its adult form. The arm is then replaced by two arms. The animal can repeat the process until it is mature.
Notable Species
Marsupitsa, Saccocoma, and Uintachrinus are considered free-swimming. Uintacrinus was a stalk-less crinoid with long arms for capturing prey. It was free-swimming the Western Interior Seaway of Cretaceous North America, 70 mya.
Pentacrinites is a fossil species which attached to driftwood. Pentacrinites evolved the ability to grow lengthy stems, even several meters long. This was likely a response to the sinking wood.
Delocrinus missouriensis is the state fossil of Missouri. It is found in Pennsylvanian period deposits.
Ancyrocrinus is found in Devonian sediments. Its cirri had evolved into a grappling hook.
Himerocrinus has been found with more than 80 arms.





